Дезоксирибонуклеи́новая молекула (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. Молекула ДНК хранит биологическую информацию в виде генетического кода, состоящего из последовательности нуклеотидов[1]. ДНК содержит информацию о структуре различных видов РНК и белков.
Структура ДНК (двойная спираль). Различные атомы в структуре показаны в разных цветах; детальная структура двух пар оснований показана снизу справаЗапрос «DNA» перенаправляется сюда; см. также другие значения. Двойная спираль (двойной винт) ДНК (правый)
Двойная спираль (двойной винт) ДНК (правый)
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органеллах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
С химической точки зрения ДНК — длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии. В целом структура молекулы ДНК получила традиционное, но ошибочное название «двойной спирали», на самом же деле она является «двойным винтом». Винтовая линия может быть правой (A- и B-формы ДНК) или левой (Z-форма ДНК)[2].
В ДНК встречается четыре вида азотистых оснований (аденин (A), гуанин (G), тимин (T) и цитозин (C)). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин (A) соединяется только с тимином (T), гуанин (G) — только с цитозином (C). Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции. Кроме того, в геноме эукариот часто встречаются участки, принадлежащие «генетическим паразитам», например транспозонам.
Расшифровка структуры ДНК (1953 год) стала одним из поворотных моментов в истории биологии. За выдающийся вклад в это открытие Фрэнсису Крику, Джеймсу Уотсону и Морису Уилкинсу была присуждена Нобелевская премия по физиологии или медицине 1962 года. Розалинд Франклин, которая получила рентгенограммы, без которых Уотсон и Крик не имели бы возможность сделать выводы о структуре ДНК, умерла в 1958 году от рака (Нобелевскую премию не дают посмертно)[3].
Содержание
- 1 История изучения
- 2 Структура молекулы
- 3 Биологические функции
- 4 Взаимодействие с белками
- 5 Генетическая рекомбинация
- 6 Эволюция метаболизма, основанного на ДНК
- 7 См. также
- 8 Примечания
- 9 Литература
- 10 Ссылки
История изучения
 Френсис Крик
Френсис Крик Джеймс Уотсон
Джеймс Уотсон Морис Уилкинс
Морис Уилкинс Розалинд Франклин
Розалинд Франклин
ДНК как химическое вещество была выделена Иоганном Фридрихом Мишером в 1869 году из остатков клеток, содержащихся в гное. Он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота[4]. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию.
До 1930-х годов считалось, что ДНК содержится только в животных клетках, а в растительных — РНК. В 1934 году в журнале «Hoppe-Seyler’s Zeitschrift fur physiologishe Chemie»[5], затем в 1935 году в «Ученых записках МГУ»[6] вышла статья советских биохимиков А. Н. Белозерского и А. Р. Кизеля в которых доказывалось присутствие ДНК в растительных клетках. В 1936 году группой Белозерского ДНК была выделена из семян и тканей бобовых, злаковых и других растений[7]. Результатом исследований этой же группы советских учёных в 1939 – 1947 годах стала первая в мировой научной литературе информация о содержании нуклеиновых кислот у различных видов бактерий.
Постепенно было доказано, что именно ДНК, а не белки, как считалось раньше, является носителем генетической информации. Одно из первых решающих доказательств принесли эксперименты Освальда Эвери, Колина Маклауда и Маклина Маккарти (1944 г.) по трансформации бактерий. Им удалось показать, что за так называемую трансформацию (приобретение болезнетворных свойств безвредной культурой в результате добавления в неё мёртвых болезнетворных бактерий) отвечает выделенная из пневмококков ДНК. Эксперимент американских учёных Алфреда Херши и Марты Чейз (эксперимент Херши — Чейз, 1952 г.) с помеченными радиоактивными изотопами белками и ДНК бактериофагов показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг[8].
Вплоть до 50-х годов XX века точное строение ДНК, как и способ передачи наследственной информации, оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены.
В результате работы группы биохимика Эрвина Чаргаффа в 1949—1951 гг. были сформулированы так называемые правила Чаргаффа. Чаргаффу и сотрудникам удалось разделить нуклеотиды ДНК при помощи бумажной хроматографии и определить точные количественные соотношения нуклеотидов разных типов. Соотношение, выявленное для аденина (А), тимина (Т), гуанина (Г) и цитозина (Ц), оказалось следующим: количество аденина равно количеству тимина, а гуанина — цитозину: А=Т, Г=Ц[9][10]. Эти правила, наряду с данными рентгеноструктурного анализа, сыграли решающую роль в расшифровке структуры ДНК.
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и правил Чаргаффа[11]. Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премией по физиологии или медицине 1962 г. Среди лауреатов не было скончавшейся к тому времени от рака Розалинд Франклин, так как премия не присуждается посмертно[12].
Интересно, что в 1957 году американцы Александер Рич, Гэри Фелзенфелд и Дэйвид Дэйвис описали нуклеиновую кислоту, составленную тремя спиралями[13]. А в 1985—1986 годах Максим Давидович Франк-Каменецкий в Москве показал, как двухспиральная ДНК складывается в так называемую H-форму, составленную уже не двумя, а тремя нитями ДНК[14][15].
Структура молекулы
Нуклеотиды
 |
 |
 |
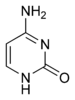 |
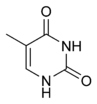
| Аденин (A) | Гуанин (G) | Тимин (T) | Цитозин (C) |
Структуры оснований в составе ДНК
Дезоксирибонуклеиновая кислота (ДНК) представляет собой биополимер (полианион), мономером которого является нуклеотид[16][17].
Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого по 5′-положению к сахару дезоксирибозе, к которому также через гликозидную связь (C—N) по 1′-положению присоединено одно из четырёх азотистых оснований.Именно наличие характерного сахара и составляет одно из главных различий между ДНК и РНК, зафиксированное в названиях этих нуклеиновых кислот (в состав РНК входит сахар рибоза)[18]. Пример нуклеотида — аденозинмонофосфат, у которого основанием, присоединённым к фосфату и рибозе, является аденин (A) (показан на рисунке).
Исходя из структуры молекул, основания, входящие в состав нуклеотидов, разделяют на две группы: пурины (аденин [A] и гуанин [G]) образованы соединёнными пяти- и шестичленным гетероциклами; пиримидины (цитозин [C] и тимин [T]) — шестичленным гетероциклом[19].
В виде исключения, например, у бактериофага PBS1, в ДНК встречается пятый тип оснований — урацил ([U]), пиримидиновое основание, отличающееся от тимина отсутствием метильной группы на кольце, обычно заменяющее тимин в РНК[20].
Следует отметить, что тимин (T) и урацил (U) не так строго приурочены к ДНК и РНК соответственно, как это считалось ранее. Так, после синтеза некоторых молекул РНК значительное число урацилов в этих молекулах метилируется с помощью специальных ферментов, превращаясь в тимин. Это происходит в транспортных и рибосомальных РНК[21].
Двойная спираль
В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке представлены формы A, B и Z (слева направо)
Полимер ДНК обладает довольно сложной структурой. Нуклеотиды соединены между собой ковалентно в длинные полинуклеотидные цепи. Эти цепи в подавляющем большинстве случаев (кроме некоторых вирусов, обладающих одноцепочечными ДНК-геномами) попарно объединяются при помощи водородных связей во вторичную структуру, получившую название двойной спирали[11][18].Остов каждой из цепей состоит из чередующихся фосфатов и сахаров[22]. Внутри одной цепи ДНК соседние нуклеотиды соединены фосфодиэфирными связями, которые формируются в результате взаимодействия между 3′-гидроксильной (3’—ОН) группой молекулы дезоксирибозы одного нуклеотида и 5′-фосфатной группой (5’—РО3) другого. Асимметричные концы цепи ДНК называются 3′ (три прайм) и 5′ (пять прайм). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3′-концу).
Как уже было сказано выше, у подавляющего большинства живых организмов ДНК состоит не из одной, а из двух полинуклеотидных цепей. Эти две длинные цепи закручены одна вокруг другой в виде двойной спирали, стабилизированной водородными связями, образующимися между обращёнными друг к другу азотистыми основаниями входящих в неё цепей. В природе эта спираль, чаще всего, правозакрученная. Направления от 3′-конца к 5′-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу).
Ширина двойной спирали составляет от 22 до 24 Å, или 2,2—2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм)[23]. Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы.
В двойной спирали различают малую (12 Å) и большую (22 Å) бороздки[24]. Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны[25].
Образование связей между основаниями
См. также: Полимеразная цепная реакция
Каждое основание на одной из цепей связывается с одним определённым основанием на второй цепи. Такое специфическое связывание называется комплементарным. Пурины комплементарны пиримидинам (то есть способны к образованию водородных связей с ними): аденин образует связи только с тимином, а цитозин — с гуанином. В двойной спирали цепочки также связаны с помощью гидрофобных взаимодействий и стэкинга, которые не зависят от последовательности оснований ДНК[26].
Комплементарность двойной спирали означает, что информация, содержащаяся в одной цепи, содержится и в другой цепи. Обратимость и специфичность взаимодействий между комплементарными парами оснований важна для репликации ДНК и всех остальных функций ДНК в живых организмах.
Так как водородные связи нековалентны, они легко разрываются и восстанавливаются. Цепочки двойной спирали могут расходиться как замок-молния под действием ферментов (хеликазы) или при высокой температуре[27]. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ — тремя водородными связями, поэтому на разрыв ГЦ требуется больше энергии. Процент ГЦ-пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки[28]. Температура плавления нуклеиновых кислот зависит от ионного окружения, рост ионной силы стабилизирует ДНК по отношению к денатурированию. При добавлении к ДНК хлорида натрия существует линейная зависимость между температурой плавления и логарифмом ионной силы раствора. Предполагается, что добавление электролита ведет к экранированию зарядов в цепях ДНК и этим уменьшает силы электростатического отталкивания между заряженными фосфатными группами, способствуя жесткости структуры. Аналогично температуру плавления ДНК повышают ионы марганца, кобальта, цинка и никеля, но ионы меди, кадмия и свинца, напротив, понижают ее[29].
Части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, ТАТА последовательность в бактериальных промоторах, обычно содержат большое количество А и Т.
Химические модификации азотистых оснований
Основная статья: Метилирование ДНК
 |
 |
 |
| Цитозин | 5-метилцитозин | Тимин |
Структура цитозина, 5-метилцитозина и тимина. Тимин может возникать путём деаминирования 5-метилцитозина
Азотистые основания в составе ДНК могут быть ковалентно модифицированы, что используется при регуляции экспрессии генов. Например, в клетках позвоночных метилирование цитозина с образованием 5-метилцитозина используется соматическими клетками для передачи профиля генной экспрессии дочерним клеткам. Метилирование цитозина не влияет на спаривание оснований в двойной спирали ДНК. У позвоночных метилирование ДНК в соматических клетках ограничивается метилированием цитозина в последовательности ЦГ[30]. Средний уровень метилирования отличается у разных организмов, так, у нематоды Caenorhabditis elegans метилирование цитозина не наблюдается, а у позвоночных обнаружен высокий уровень метилирования — до 1 %[31]. Другие модификации оснований включают метилирование аденина у бактерий и гликозилирование урацила с образованием «J-основания» в кинетопластах[32].
Метилирование цитозина с образованием 5-метилцитозина в промоторной части гена коррелирует с его неактивным состоянием[33]. Метилирование цитозина важно также для инактивации Х-хромосомы у млекопитающих[34]. Метилирование ДНК используется в геномном импринтинге[35]. Значительные нарушения профиля метилирования ДНК происходят при канцерогенезе[36].
Несмотря на биологическую роль, 5-метилцитозин может спонтанно утрачивать аминную группу (деаминироваться), превращаясь в тимин, поэтому метилированные цитозины являются источником повышенного числа мутаций[37].
Повреждения ДНК
Основная статья: Мутация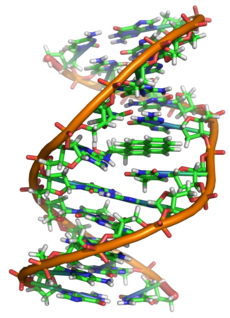 Интеркалированное химическое соединение, которое находится в середине спирали — бензопирен, основной мутаген табачного дыма[38]
Интеркалированное химическое соединение, которое находится в середине спирали — бензопирен, основной мутаген табачного дыма[38]
ДНК может повреждаться разнообразными мутагенами, к которым относятся окисляющие и алкилирующие вещества, а также высокоэнергетическая электромагнитная радиация — ультрафиолетовое и рентгеновское излучение. Тип повреждения ДНК зависит от типа мутагена. Например, ультрафиолет повреждает ДНК путём образования в ней димеров тимина, которые возникают при образовании ковалентных связей между соседними основаниями[39].
Оксиданты, такие как свободные радикалы или пероксид водорода, приводят к нескольким типам повреждения ДНК, включая модификации оснований, в особенности гуанозина, а также двухцепочечные разрывы в ДНК[40]. По некоторым оценкам, в каждой клетке человека окисляющими соединениями ежедневно повреждается порядка 500 оснований[41][42]. Среди разных типов повреждений наиболее опасные — это двухцепочечные разрывы, потому что они трудно репарируются и могут привести к потерям участков хромосом (делециям) и транслокациям.
Многие молекулы мутагенов вставляются (интеркалируют) между двумя соседними парами оснований. Большинство этих соединений, например: бромистый этидий, даунорубицин, доксорубицин и талидомид, имеет ароматическую структуру. Для того чтобы интеркалирующее соединение могло поместиться между основаниями, они должны разойтись, расплетая и нарушая структуру двойной спирали. Эти изменения в структуре ДНК мешают транскрипции и репликации, вызывая мутации. Поэтому интеркалирующие соединения часто являются канцерогенами, наиболее известные из которых — бензопирен, акридины, афлатоксин и бромистый этидий[43][44][45]. Несмотря на эти негативные свойства, в силу их способности подавлять транскрипцию и репликацию ДНК, интеркалирующие соединения используются в химиотерапии для подавления быстро растущих клеток рака[46].
Некоторые вещества (цисплатин[47], митомицин C[48], псорален[49]) образуют поперечные сшивки между нитями ДНК и подавляют синтез ДНК, благодаря чему используются в химиотерапии некоторых видов рака (см. Химиотерапия злокачественных новообразований).
Суперскрученность
Основная статья: Суперскрученная ДНК
Если взяться за концы верёвки и начать скручивать их в разные стороны, она становится короче и на верёвке образуются «супервитки». Так же может быть суперскручена и ДНК. В обычном состоянии цепочка ДНК делает один оборот на каждые 10,4 пар оснований, но в суперскрученном состоянии спираль может быть свёрнута туже или расплетена[50]. Выделяют два типа суперскручивания: положительное — в направлении нормальных витков, при котором основания расположены ближе друг к другу; и отрицательное — в противоположном направлении. В природе молекулы ДНК обычно находятся в отрицательном суперскручивании, которое вносится ферментами — топоизомеразами[51]. Эти ферменты удаляют дополнительное скручивание, возникающее в ДНК в результате транскрипции и репликации[52].
 Структура теломер. Зелёным цветом показан ион металла, хелатированный в центре структуры[53]
Структура теломер. Зелёным цветом показан ион металла, хелатированный в центре структуры[53]
Структуры на концах хромосом
На концах линейных хромосом находятся специализированные структуры ДНК, называемые теломерами. Основная функция этих участков — поддержание целостности концов хромосом[54]. Теломеры также защищают концы ДНК от деградации экзонуклеазами и предотвращают активацию системы репарации[55]. Поскольку обычные ДНК-полимеразы не могут реплицировать 3′ концы хромосом, это делает специальный фермент — теломераза.
В клетках человека теломеры часто представлены одноцепочечной ДНК и состоят из нескольких тысяч повторяющихся единиц последовательности ТТАГГГ[56]. Эти последовательности с высоким содержанием гуанина стабилизируют концы хромосом, формируя очень необычные структуры, называемые G-квадруплексами и состоящие из четырёх, а не двух взаимодействующих оснований. Четыре гуаниновых основания, все атомы которых находятся в одной плоскости, образуют пластинку, стабилизированную водородными связями между основаниями и хелатированием в центре неё иона металла (чаще всего калия). Эти пластинки располагаются стопкой друг над другом[57].
На концах хромосом могут образовываться и другие структуры: основания могут быть расположены в одной цепочке или в разных параллельных цепочках. Кроме этих «стопочных» структур теломеры формируют большие петлеобразные структуры, называемые Т-петли или теломерные петли. В них одноцепочечная ДНК располагается в виде широкого кольца, стабилизированного теломерными белками[58]. В конце Т-петли одноцепочечная теломерная ДНК присоединяется к двухцепочечной ДНК, нарушая спаривание цепочек в этой молекуле и образуя связи с одной из цепей. Это трёхцепочечное образование называется Д-петля (от англ. displacement loop)[57].
Биологические функции
ДНК является носителем генетической информации, записанной в виде последовательности нуклеотидов с помощью генетического кода. С молекулами ДНК связаны два основополагающих свойства живых организмов — наследственность и изменчивость. В ходе процесса, называемого репликацией ДНК, образуются две копии исходной цепочки, наследуемые дочерними клетками при делении, отсюда следует, что образовавшиеся клетки оказываются генетически идентичны исходной.
Генетическая информация реализуется при экспрессии генов в процессах транскрипции (синтеза молекул РНК на матрице ДНК) и трансляции (синтеза белков на матрице РНК).
Последовательность нуклеотидов «кодирует» информацию о различных типах РНК: информационных, или матричных (мРНК), рибосомальных (рРНК) и транспортных (тРНК). Все эти типы РНК синтезируются на основе ДНК в процессе транскрипции. Роль их в биосинтезе белков (процессе трансляции) различна. Информационная РНК содержит информацию о последовательности аминокислот в белке, рибосомальные РНК служат основой для рибосом (сложных нуклеопротеиновых комплексов, основная функция которых — сборка белка из отдельных аминокислот на основе иРНК), транспортные РНК доставляют аминокислоты к месту сборки белков — в активный центр рибосомы, «ползущей» по иРНК.
Структура генома
Основные статьи: Геном, Ген, Клеточное ядро, Хроматин и Хромосома ДНК генома бактериофага: фотография под просвечивающим электронным микроскопом
ДНК генома бактериофага: фотография под просвечивающим электронным микроскопом
Большинство природных ДНК имеет двухцепочечную структуру, линейную (эукариоты, некоторые вирусы и отдельные роды бактерий) или кольцевую (прокариоты, хлоропласты и митохондрии). Линейную одноцепочечную ДНК содержат некоторые вирусы и бактериофаги.Молекулы ДНК находятся in vivo в плотно упакованном, конденсированном состоянии[59]. В клетках эукариот ДНК располагается главным образом в ядре и на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа в виде набора хромосом. Бактериальная (прокариоты) ДНК обычно представлена одной кольцевой молекулой ДНК, расположенной в неправильной формы образовании в цитоплазме, называемым нуклеоидом[60]. Генетическая информация генома состоит из генов. Ген — единица передачи наследственной информации и участок ДНК, который влияет на определённую характеристику организма. Ген содержит открытую рамку считывания, которая транскрибируется, а также регуляторные последовательности (англ.) (рус., например промотор и энхансер, которые контролируют экспрессию открытых рамок считывания.
У многих видов только малая часть общей последовательности генома кодирует белки. Так, только около 1,5 % генома человека состоит из кодирующих белок экзонов, а больше 50 % ДНК человека состоит из некодирующих повторяющихся последовательностей ДНК[61]. Причины наличия такого большого количества некодирующей ДНК в эукариотических геномах и огромная разница в размерах геномов (С-значение) — одна из неразрешённых научных загадок[62]; исследования в этой области также указывают на большое количество фрагментов реликтовых вирусов в этой части ДНК.
Последовательности генома, не кодирующие белок
Основная статья: Некодирующая ДНК
В настоящее время накапливается всё больше данных, противоречащих идее о некодирующих последовательностях как «мусорной ДНК» (англ. junk DNA).Теломеры и центромеры содержат малое число генов, но они важны для функционирования и стабильности хромосом[55][63]. Часто встречающаяся форма некодирующих последовательностей человека — псевдогены, копии генов, инактивированные в результате мутаций[64]. Эти последовательности нечто вроде молекулярных ископаемых, хотя иногда они могут служить исходным материалом для дупликации и последующей дивергенции генов[65].Другой источник разнообразия белков в организме — это использование интронов в качестве «линий разреза и склеивания» в альтернативном сплайсинге[66].Наконец, не кодирующие белок последовательности могут кодировать вспомогательные клеточные РНК, например мяРНК[67]. Недавнее исследование транскрипции генома человека показало, что 10 % генома даёт начало полиаденилированным РНК[68], а исследование генома мыши показало, что 62 % его транскрибируется[69].
Транскрипция и трансляция
Основные статьи: Генетический код, Транскрипция (биология) и Трансляция (биология)
Генетическая информация, закодированная в ДНК, должна быть прочитана и в конечном итоге выражена в синтезе различных биополимеров, из которых состоят клетки. Последовательность оснований в цепочке ДНК напрямую определяет последовательность оснований в РНК, на которую она «переписывается» в процессе, называемом транскрипцией. В случае мРНК эта последовательность определяет аминокислоты белка. Соотношение между нуклеотидной последовательностью мРНК и аминокислотной последовательностью определяется правилами трансляции, которые называются генетическим кодом. Генетический код состоит из трёхбуквенных «слов», называемых кодонами, состоящих из трёх нуклеотидов (то есть ACT, CAG, TTT и т. п.).Во время транскрипции нуклеотиды гена копируются на синтезируемую РНК РНК-полимеразой. Эта копия в случае мРНК декодируется рибосомой, которая «читает» последовательность мРНК, осуществляя спаривание матричной РНК с транспортными РНК, которые присоединены к аминокислотам. Поскольку в трёхбуквенных комбинациях используются 4 основания, всего возможны 64 кодона (4³ комбинации). Кодоны кодируют 20 стандартных аминокислот, каждой из которых соответствует в большинстве случаев более одного кодона. Один из трёх кодонов, которые располагаются в конце мРНК, не означает аминокислоту и определяет конец белка, это «стоп» или «нонсенс» кодоны — TAA, TGA, TAG.
Репликация
Основная статья: Репликация ДНК
Деление клеток необходимо для размножения одноклеточного и роста многоклеточного организма, но до деления клетка должна удвоить геном, чтобы дочерние клетки содержали ту же генетическую информацию, что и исходная клетка. Из нескольких теоретически возможных механизмов удвоения (репликации) ДНК реализуется полуконсервативный. Две цепочки разделяются, а затем каждая недостающая комплементарная последовательность ДНК воспроизводится ферментом ДНК-полимеразой. Этот фермент синтезирует полинуклеотидную цепь, находя правильный нуклеотид через комплементарное спаривание оснований и присоединяя его к растущей цепочке. ДНК-полимераза не может начинать новую цепь, а может лишь наращивать уже существующую, поэтому она нуждается в короткой цепочке нуклеотидов — (праймере), синтезируемом праймазой. Так как ДНК-полимеразы могут синтезировать цепочку только в направлении 5′ —> 3′, антипараллельные цепи ДНК копируются по-разному: одна цепь синтезируется непрерывно, а вторая прерывчато[70].
Взаимодействие с белками
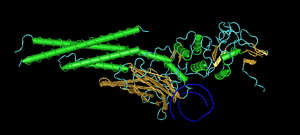 Взаимодействие фактора транскрипции STAT3 с ДНК (показана в виде синей спирали)
Взаимодействие фактора транскрипции STAT3 с ДНК (показана в виде синей спирали)
Все функции ДНК зависят от её взаимодействия с белками. Взаимодействия могут быть неспецифическими, когда белок присоединяется к любой молекуле ДНК, или зависеть от наличия особой последовательности. Ферменты также могут взаимодействовать с ДНК, из них наиболее важные — это РНК-полимеразы, которые копируют последовательность оснований ДНК на РНК в транскрипции или при синтезе новой цепи ДНК — репликации.
Структурные и регуляторные белки
Хорошо изученными примерами взаимодействия белков и ДНК, не зависящего от нуклеотидной последовательности ДНК, является взаимодействие со структурными белками. В клетке ДНК связана с этими белками, образуя компактную структуру, которая называется хроматин. У эукариот хроматин образован при присоединении к ДНК небольших щелочных белков — гистонов, менее упорядоченный хроматин прокариот содержит гистон-подобные белки[71][72]. Гистоны формируют дискообразную белковую структуру — нуклеосому, вокруг каждой из которых вмещается два оборота спирали ДНК. Неспецифические связи между гистонами и ДНК образуются за счёт ионных связей щелочных аминокислот гистонов и кислотных остатков сахарофосфатного остова ДНК[73]. Химические модификации этих аминокислот включают метилирование, фосфорилирование и ацетилирование[74]. Эти химические модификации изменяют силу взаимодействия между ДНК и гистонами, влияя на доступность специфических последовательностей для факторов транскрипции и изменяя скорость транскрипции[75]. Другие белки в составе хроматина, которые присоединяются к неспецифическим последовательностям — белки с высокой подвижностью в гелях, которые ассоциируют большей частью с согнутой ДНК[76]. Эти белки важны для образования в хроматине структур более высокого порядка[77].
Особая группа белков, присоединяющихся к ДНК — это белки, которые ассоциируют с одноцепочечной ДНК. Наиболее хорошо охарактеризованный белок этой группы у человека — репликационный белок А, без которого невозможно протекание большинства процессов, где расплетается двойная спираль, включая репликацию, рекомбинацию и репарацию. Белки этой группы стабилизируют одноцепочечную ДНК и предотвращают формирование стеблей-петель или деградации нуклеазами[78].
В то же время другие белки узнают и присоединяются к специфическим последовательностям. Наиболее изученная группа таких белков — различные классы факторов транскрипции, то есть белки, регулирующие транскрипцию. Каждый из этих белков узнаёт свою последовательность, часто в промоторе, и активирует или подавляет транскрипцию гена. Это происходит при ассоциации факторов транскрипции с РНК-полимеразой либо напрямую, либо через белки-посредники. Полимераза ассоциирует сначала с белками, а потом начинает транскрипцию[79]. В других случаях факторы транскрипции могут присоединяться к ферментам, которые модифицируют находящиеся на промоторах гистоны, что изменяет доступность ДНК для полимераз[80].
Так как специфические последовательности встречаются во многих местах генома, изменения в активности одного типа фактора транскрипции могут изменить активность тысяч генов[81]. Соответственно, эти белки часто регулируются в процессах ответа на изменения в окружающей среде, развития организма и дифференцировки клеток. Специфичность взаимодействия факторов транскрипции с ДНК обеспечивается многочисленными контактами между аминокислотами и основаниями ДНК, что позволяет им «читать» последовательность ДНК. Большинство контактов с основаниями происходит в главной бороздке, где основания более доступны[25].
Ферменты, модифицирующие ДНК
Топоизомеразы и хеликазы
Основные статьи: Топоизомеразы и Хеликазы
В клетке ДНК находится в компактном, т. н. суперскрученном состоянии, иначе она не смогла бы в ней уместиться. Для протекания жизненно важных процессов ДНК должна быть раскручена, что производится двумя группами белков — топоизомеразами и хеликазами.
Топоизомеразы — ферменты, которые имеют и нуклеазную, и лигазную активности. Они изменяют степень суперскрученности в ДНК. Некоторые из этих ферментов разрезают спираль ДНК и позволяют вращаться одной из цепей, тем самым уменьшая уровень суперскрученности, после чего фермент заделывает разрыв[51]. Другие ферменты могут разрезать одну из цепей и проводить вторую цепь через разрыв, а потом лигировать разрыв в первой цепи[82]. Топоизомеразы необходимы во многих процессах, связанных с ДНК, таких как репликация и транскрипция[52].
Хеликазы — белки, которые являются одним из молекулярных моторов. Они используют химическую энергию нуклеотидтрифосфатов, чаще всего АТФ, для разрыва водородных связей между основаниями, раскручивая двойную спираль на отдельные цепочки[83]. Эти ферменты важны для большинства процессов, где белкам необходим доступ к основаниям ДНК.
Нуклеазы и лигазы
Основные статьи: Нуклеаза и Лигаза
В различных процессах, происходящих в клетке, например рекомбинации и репарации, участвуют ферменты, способные разрезать и восстанавливать целостность нитей ДНК. Ферменты, разрезающие ДНК, носят название нуклеаз. Нуклеазы, которые гидролизуют нуклеотиды на концах молекулы ДНК, называются экзонуклеазами, а эндонуклеазы разрезают ДНК внутри цепи. Наиболее часто используемые в молекулярной биологии и генетической инженерии нуклеазы — это эндонуклеазы рестрикции (рестриктазы), которые разрезают ДНК около специфических последовательностей. Например, фермент EcoRV (рестрикционный фермент № 5 из ‘E. coli‘) узнаёт шестинуклеотидную последовательность 5′-GAT|ATC-3′ и разрезает ДНК в месте, указанном вертикальной линией. В природе эти ферменты защищают бактерии от заражения бактериофагами, разрезая ДНК фага, когда она вводится в бактериальную клетку. В этом случае нуклеазы — часть системы модификации-рестрикции[84]. ДНК-лигазы «сшивают» концы фрагментов ДНК между собой, катализируя формирование фосфодиэфирной связи с использованием энергии АТФ.Рестрикционные нуклеазы и лигазы используются в клонировании и фингерпринтинге.
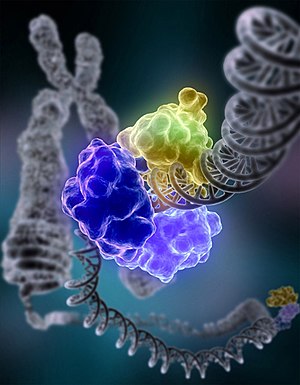 ДНК-лигаза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных разными цветами), лигирующая повреждённую цепь ДНК
ДНК-лигаза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных разными цветами), лигирующая повреждённую цепь ДНК
Полимеразы
Основная статья: ДНК-полимераза
Существует также важная для метаболизма ДНК группа ферментов, которые синтезируют цепи полинуклеотидов из нуклеозидтрифосфатов — ДНК-полимеразы. Они добавляют нуклеотиды к 3′-гидроксильной группе предыдущего нуклеотида в цепи ДНК, поэтому все полимеразы работают в направлении 5′—> 3′[85]. В активном центре этих ферментов субстрат — нуклеозидтрифосфат — спаривается с комплементарным основанием в составе одноцепочечной полинуклеотидной цепочки — матрицы.
В процессе репликации ДНК ДНК-зависимая ДНК-полимераза синтезирует копию исходной последовательности ДНК. Точность очень важна в этом процессе, так как ошибки в полимеризации приведут к мутациям, поэтому многие полимеразы обладают способностью к «редактированию» — исправлению ошибок. Полимераза узнаёт ошибки в синтезе по отсутствию спаривания между неправильными нуклеотидами. После определения отсутствия спаривания активируется 3′—> 5′ экзонуклеазная активность полимеразы, и неправильное основание удаляется[86]. В большинстве организмов ДНК-полимеразы работают в виде большого комплекса, называемого реплисомой, которая содержит многочисленные дополнительные субъединицы, например хеликазы[87].
РНК-зависимые ДНК-полимеразы — специализированный тип полимераз, которые копируют последовательность РНК на ДНК. К этому типу относятся обратная транскриптаза, которая содержится в ретровирусах и используется при инфекции клеток, а также теломераза, необходимая для репликации теломер[88]. Теломераза — необычный фермент, потому что она содержит собственную матричную РНК[55].
Транскрипция осуществляется ДНК-зависимой РНК-полимеразой, которая копирует последовательность ДНК одной цепочки на мРНК. В начале транскрипции гена РНК-полимераза присоединяется к последовательности в начале гена, называемой промотором, и расплетает спираль ДНК. Потом она копирует последовательность гена на матричную РНК до тех пор, пока не дойдёт до участка ДНК в конце гена — терминатора, где она останавливается и отсоединяется от ДНК. Также как ДНК-зависимая ДНК-полимераза человека, РНК-полимераза II, которая транскрибирует большую часть генов в геноме человека, работает в составе большого белкового комплекса, содержащего регуляторные и дополнительные единицы[89].
Генетическая рекомбинация
Основная статья: Рекомбинация (биология) Рекомбинация происходит в результате физического разрыва в хромосомах (М) и (F) и их последующего соединения с образованием двух новых хромосом (C1 и C2)
Рекомбинация происходит в результате физического разрыва в хромосомах (М) и (F) и их последующего соединения с образованием двух новых хромосом (C1 и C2)
Двойная спираль ДНК обычно не взаимодействует с другими сегментами ДНК, и в человеческих клетках разные хромосомы пространственно разделены в ядре[90]. Это расстояние между разными хромосомами важно для способности ДНК действовать в качестве стабильного носителя информации. В процессе рекомбинации с помощью ферментов две спирали ДНК разрываются, обмениваются участками, после чего непрерывность спиралей восстанавливается, поэтому обмен участками негомологичных хромосом может привести к повреждению целостности генетического материала.
Рекомбинация позволяет хромосомам обмениваться генетической информацией, в результате этого образуются новые комбинации генов, что увеличивает эффективность естественного отбора и важно для быстрой эволюции новых белков[91]. Генетическая рекомбинация также играет роль в репарации, особенно в ответе клетки на разрыв обеих цепей ДНК[92].
Самая распространённая форма кроссинговера — это гомологичная рекомбинация, когда принимающие участие в рекомбинации хромосомы имеют очень похожие последовательности. Иногда в качестве участков гомологии выступают транспозоны. Негомологичная рекомбинация может привести к повреждению клетки, поскольку в результате такой рекомбинации возникают транслокации. Реакция рекомбинации катализируется ферментами, которые называются рекомбиназы, например, Cre. На первом этапе реакции рекомбиназа делает разрыв в одной из цепей ДНК, позволяя этой цепи отделиться от комплементарной цепи и присоединиться к одной из цепей второй хроматиды. Второй разрыв в цепи второй хроматиды позволяет ей также отделиться и присоединиться к оставшейся без пары цепи из первой хроматиды, формируя структуру Холлидея. Структура Холлидея может передвигаться вдоль соединённой пары хромосом, меняя цепи местами. Реакция рекомбинации завершается, когда фермент разрезает соединение, а две цепи лигируются[93].
Эволюция метаболизма, основанного на ДНК
ДНК содержит генетическую информацию, которая делает возможной жизнедеятельность, рост, развитие и размножение всех современных организмов. Однако как долго в течение четырёх миллиардов лет истории жизни на Земле ДНК была главным носителем генетической информации, неизвестно. Существуют гипотезы, что РНК играла центральную роль в обмене веществ, поскольку она может и переносить генетическую информацию, и осуществлять катализ с помощью рибозимов[94][95][96]. Кроме того, РНК — один из основных компонентов «фабрик белка» — рибосом. Древний РНК-мир, где нуклеиновая кислота была использована и для катализа, и для переноса информации, мог послужить источником современного генетического кода, состоящего из четырёх оснований. Это могло произойти в результате того, что число оснований в организме было компромиссом между небольшим числом оснований, увеличивавшим точность репликации, и большим числом оснований, увеличивающим каталитическую активность рибозимов[97].
К сожалению, древние генетические системы не дошли до наших дней. ДНК в окружающей среде в среднем сохраняется в течение 1 миллиона лет, постепенно деградируя до коротких фрагментов. Извлечение ДНК из бактериальных спор, заключённых в кристаллах соли 250 млн лет назад, и определение последовательности генов 16S рРНК[98], служит темой оживлённой дискуссии в научной среде[99][100].
См. также
- Вектор (биология)
- Геном человека
- Действие излучений на структуру и функции ДНК
- Методы секвенирования нового поколения
- Мобильные элементы генома
- Нуклеопротеиды
- Спиртовая преципитация
- Футпринтинг ДНК
- Центральная догма молекулярной биологии
- Цис-элемент
- ДНК-компьютер
Примечания
- ↑ Александр Панчин. Сумма биотехнологии [1]. — АСТ, 2015. — С. 13. — 432 с. — ISBN 978-5-17-093602-1.
- ↑ Bustamante C., Bryant Z., Smith S. B. Ten years of tension: single-molecule DNA mechanics (англ.) // Nature. — 2003. — Vol. 421, no. 6921. — P. 423—427.
- ↑ Erica Westly. No Nobel for You: Top 10 Nobel Snubs. Rosalind Franklin—her work on the structure of DNA never received a Nobel (англ.). Scientific American (6 октября 2008). Дата обращения: 18 ноября 2013. Архивировано 8 января 2014 года.
- ↑ Dahm R. Friedrich Miescher and the discovery of DNA (англ.) // Dev Biol (англ.) (рус. : journal. — 2005. — Vol. 278, no. 2. — P. 274—288. — PMID 15680349.
- ↑ Kiesel A., Beloserskii A. Hoppe-Seyler’s Zeitschrift fur physiologishe Chemie, 229, 160—166. 1934.
- ↑ Белозерский А. Н. Ученые записки МГУ, вып.4, 209—215, 1935.
- ↑ Белозерский А. Н., Чигирев С. Д. Биохимия, 1, 136—146, 1936.
- ↑ Hershey A., Chase M. Independent functions of viral protein and nucleic acid in growth of bacteriophage (англ.) // The Journal of General Physiology (англ.) (рус. : journal. — Rockefeller University Press (англ.) (рус., 1952. — Vol. 36, no. 1. — P. 39—56. — PMID 12981234.
- ↑ Elson D., Chargaff E. On the deoxyribonucleic acid content of sea urchin gametes (англ.) // Experientia : journal. — 1952. — Vol. 8, no. 4. — P. 143—145. — doi:10.1007/BF02170221. — PMID 14945441.
- ↑ Chargaff E., Lipshitz R., Green C. Composition of the deoxypentose nucleic acids of four genera of sea-urchin (англ.) // J Biol Chem : journal. — 1952. — Vol. 195, no. 1. — P. 155—160. — PMID 14938364.
- ↑ 1 2 Watson J., Crick F. Molecular structure of nucleic acids; a structure for deoxyribose nucleic acid (рум.) // Nature. — 1953. — Т. 171, nr. 4356. — P. 737 — 8.
- ↑ The Nobel Prize in Physiology or Medicine 1962 Nobelprize .org Accessed 22 Dec 06
- ↑ Н. Домрина В России есть кому делать науку — если будет на что // Журнал «Наука и жизнь», № 2, 2002
- ↑ Maxim Frank-Kamenetskii DNA structure: A simple solution to the stability of the double helix? // Журнал Nature № 324, 305 (27 November 1986)
- ↑ Maxim Frank-Kamenetskii H-form DNA and the hairpin-triplex model // Журнал Nature № 333, 214 (19 May 1988)
- ↑ Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters. Molecular Biology of the Cell; Fourth Edition (англ.). — New York and London: Garland Science (англ.) (рус., 2002.
- ↑ Butler, John M. (2001) Forensic DNA Typing «Elsevier». pp. 14 — 15. ISBN 978-0-12-147951-0
- ↑ 1 2 Berg J., Tymoczko J. and Stryer L. (2002) Biochemistry. W. H. Freeman and Company ISBN 0-7167-4955-6
- ↑ Abbreviations and Symbols for Nucleic Acids, Polynucleotides and their Constituents IUPAC-IUB Commission on Biochemical Nomenclature (CBN) Accessed 03 Jan 2006
- ↑ Takahashi I., Marmur J. Replacement of thymidylic acid by deoxyuridylic acid in the deoxyribonucleic acid of a transducing phage for Bacillus subtilis (англ.) // Nature : journal. — 1963. — Vol. 197. — P. 794 — 5.
- ↑ Agris P. Decoding the genome: a modified view (англ.) // Nucleic Acids Res (англ.) (рус. : journal. — 2004. — Vol. 32, no. 1. — P. 223 — 38. — PMID 14715921.
- ↑ Ghosh A., Bansal M. A glossary of DNA structures from A to Z (англ.) // Acta Crystallogr D Biol Crystallogr (англ.) (рус. : journal. — International Union of Crystallography, 2003. — Vol. 59, no. Pt 4. — P. 620 — 6.
- ↑ Mandelkern M., Elias J., Eden D., Crothers D. The dimensions of DNA in solution (англ.) // J Mol Biol (англ.) (рус. : journal. — 1981. — Vol. 152, no. 1. — P. 153 — 61.
- ↑ Wing R., Drew H., Takano T., Broka C., Tanaka S., Itakura K., Dickerson R. Crystal structure analysis of a complete turn of B-DNA (англ.) // Nature : journal. — 1980. — Vol. 287, no. 5784. — P. 755 — 8.
- ↑ 1 2 Pabo C., Sauer R. Protein-DNA recognition (англ.) // Annu Rev Biochem (англ.) (рус. : journal. — Vol. 53. — P. 293 — 321.
- ↑ Ponnuswamy P., Gromiha M. On the conformational stability of oligonucleotide duplexes and tRNA molecules (англ.) // J Theor Biol (англ.) (рус. : journal. — 1994. — Vol. 169, no. 4. — P. 419—432. — PMID 7526075.
- ↑ Clausen-Schaumann H., Rief M., Tolksdorf C., Gaub H. Mechanical stability of single DNA molecules (англ.) // Biophys J (англ.) (рус. : journal. — 2000. — Vol. 78, no. 4. — P. 1997—2007. — PMID 10733978.
- ↑ Chalikian T., Völker J., Plum G., Breslauer K. A more unified picture for the thermodynamics of nucleic acid duplex melting: a characterization by calorimetric and volumetric techniques (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1999. — Vol. 96, no. 14. — P. 7853—7858. — PMID 10393911.
- ↑ Е.Е.Крисс, К.Б.Яцимирский. Взаимодействие нуклеиновых кислот с металлами..
- ↑ Молекулярная биология клетки: в 3-х томах / Б. Альбертс, А. Джонсон, Д. Льюис и др. — М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. — Т. I. — С. 719—733. — 808 с. — ISBN 978-5-4344-0112-8.
- ↑ Bird A. DNA methylation patterns and epigenetic memory (англ.) // Genes Dev : journal. — 2002. — Vol. 16, no. 1. — P. 6 — 21.
- ↑ Gommers-Ampt J., Van Leeuwen F., de Beer A., Vliegenthart J., Dizdaroglu M., Kowalak J., Crain P., Borst P. beta-D-glucosyl-hydroxymethyluracil: a novel modified base present in the DNA of the parasitic protozoan T. brucei (англ.) // Cell : journal. — Cell Press, 1993. — Vol. 75, no. 6. — P. 1129 — 36.
- ↑ Jones P. A. Functions of DNA methylation: islands, start sites, gene bodies and beyond // Nature Reviews Genetics. — 2012. — Т. 13, № 7. — С. 484—492.
- ↑ Klose R., Bird A. Genomic DNA methylation: the mark and its mediators (англ.) // Trends Biochem Sci (англ.) (рус. : journal. — 2006. — Vol. 31, no. 2. — P. 89 — 97.
- ↑ Li E., Beard C., Jaenisch R. Role for DNA methylation in genomic imprinting //Nature. — 1993. — Т. 366. — №. 6453. — С. 362—365
- ↑ Ehrlich M. DNA methylation in cancer: too much, but also too little //Oncogene. — 2002. — Т. 21. — №. 35. — С. 5400-5413
- ↑ Walsh C., Xu G. Cytosine methylation and DNA repair (неопр.) // Curr Top Microbiol Immunol. — Т. 301. — С. 283 — 315.
- ↑ Created from PDB 1JDG
- ↑ Douki T., Reynaud-Angelin A., Cadet J., Sage E. Bipyrimidine photoproducts rather than oxidative lesions are the main type of DNA damage involved in the genotoxic effect of solar UVA radiation (англ.) // Biochemistry : journal. — 2003. — Vol. 42, no. 30. — P. 9221 — 6.
- ↑ Cadet J., Delatour T., Douki T., Gasparutto D., Pouget J., Ravanat J., Sauvaigo S. Hydroxyl radicals and DNA base damage (неопр.) // Mutation Research (англ.) (рус.. — Elsevier, 1999. — Т. 424, № 1 — 2. — С. 9 — 21.
- ↑ Shigenaga M., Gimeno C., Ames B. Urinary 8-hydroxy-2′-deoxyguanosine as a biological marker of in vivo oxidative DNA damage (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1989. — Vol. 86, no. 24. — P. 9697 — 701.
- ↑ Cathcart R., Schwiers E., Saul R., Ames B. Thymine glycol and thymidine glycol in human and rat urine: a possible assay for oxidative DNA damage (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1984. — Vol. 81, no. 18. — P. 5633 — 7.
- ↑ Ferguson L., Denny W. The genetic toxicology of acridines (неопр.) // Mutation Research (англ.) (рус.. — Elsevier, 1991. — Т. 258, № 2. — С. 123 — 60.
- ↑ Jeffrey A. DNA modification by chemical carcinogens (англ.) // Pharmacol Ther : journal. — 1985. — Vol. 28, no. 2. — P. 237 — 72.
- ↑ Stephens T., Bunde C., Fillmore B. Mechanism of action in thalidomide teratogenesis (англ.) // Biochem Pharmacol (англ.) (рус. : journal. — 2000. — Vol. 59, no. 12. — P. 1489 — 99.
- ↑ Braña M., Cacho M., Gradillas A., de Pascual-Teresa B., Ramos A. Intercalators as anticancer drugs (англ.) // Curr Pharm Des (англ.) (рус. : journal. — 2001. — Vol. 7, no. 17. — P. 1745 — 80.
- ↑ Trzaska, Stephen. Cisplatin (англ.) // Chemical & Engineering News (англ.) (рус. : journal. — 2005. — 20 June (vol. 83, no. 25).
- ↑ Tomasz, Maria. Mitomycin C: small, fast and deadly (but very selective) (англ.) // Chemistry and Biology (англ.) (рус. : journal. — 1995. — September (vol. 2, no. 9). — P. 575—579. — doi:10.1016/1074-5521(95)90120-5. — PMID 9383461.
- ↑ Wu Q., Christensen L. A., Legerski R. J., Vasquez K. M. Mismatch repair participates in error-free processing of DNA interstrand crosslinks in human cells (англ.) // EMBO Rep. (англ.) (рус. : journal. — 2005. — June (vol. 6, no. 6). — P. 551—557. — doi:10.1038/sj.embor.7400418. — PMID 15891767.
- ↑ Benham C., Mielke S. DNA mechanics (неопр.) // Annu Rev Biomed Eng (англ.) (рус.. — 2005. — Т. 7. — С. 21—53. — PMID 16004565.
- ↑ 1 2 Champoux J. DNA topoisomerases: structure, function, and mechanism (англ.) // Annu Rev Biochem (англ.) (рус. : journal. — 2001. — Vol. 70. — P. 369—413. — PMID 11395412.
- ↑ 1 2 Wang J. Cellular roles of DNA topoisomerases: a molecular perspective (англ.) // Nat Rev Mol Cell Biol : journal. — 2002. — Vol. 3, no. 6. — P. 430—440. — PMID 12042765.
- ↑ Created from NDB UD0017 Архивировано 7 июня 2013 года.
- ↑ Greider C., Blackburn E. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts (англ.) // Cell : journal. — Cell Press, 1985. — Vol. 43, no. 2 Pt 1. — P. 405—413. — PMID 3907856.
- ↑ 1 2 3 Nugent C., Lundblad V. The telomerase reverse transcriptase: components and regulation (англ.) // Genes Dev : journal. — 1998. — Vol. 12, no. 8. — P. 1073—1085. — PMID 9553037.
- ↑ Wright W., Tesmer V., Huffman K., Levene S., Shay J. Normal human chromosomes have long G-rich telomeric overhangs at one end (англ.) // Genes Dev : journal. — 1997. — Vol. 11, no. 21. — P. 2801—2809. — PMID 9353250.
- ↑ 1 2 Burge S., Parkinson G., Hazel P., Todd A., Neidle S. Quadruplex DNA: sequence, topology and structure (англ.) // Nucleic Acids Res (англ.) (рус. : journal. — 2006. — Vol. 34, no. 19. — P. 5402—5415. — PMID 17012276.
- ↑ Griffith J., Comeau L., Rosenfield S., Stansel R., Bianchi A., Moss H., de Lange T. Mammalian telomeres end in a large duplex loop (англ.) // Cell. — Cell Press, 1999. — Vol. 97, no. 4. — P. 503—514. — PMID 10338214.
- ↑ Teif V.B. and Bohinc K. Condensed DNA: condensing the concepts (неопр.) // Progress in Biophysics and Molecular Biology. — 2010. — doi:10.1016/j.pbiomolbio.2010.07.002.
- ↑ Thanbichler M., Wang S., Shapiro L. The bacterial nucleoid: a highly organized and dynamic structure (англ.) // J Cell Biochem (англ.) (рус. : journal. — 2005. — Vol. 96, no. 3. — P. 506 — 21.
- ↑ Wolfsberg T., McEntyre J., Schuler G. Guide to the draft human genome (англ.) // Nature. — 2001. — Vol. 409, no. 6822. — P. 824 — 6.
- ↑ Gregory T. The C-value enigma in plants and animals: a review of parallels and an appeal for partnership (англ.) // Ann Bot (Lond) : journal. — 2005. — Vol. 95, no. 1. — P. 133 — 46.
- ↑ Pidoux A., Allshire R. The role of heterochromatin in centromere function (англ.) // Philos Trans R Soc Lond B Biol Sci : journal. — 2005. — Vol. 360, no. 1455. — P. 569 — 79. (недоступная ссылка)
- ↑ Harrison P., Hegyi H., Balasubramanian S., Luscombe N., Bertone P., Echols N., Johnson T., Gerstein M. Molecular fossils in the human genome: identification and analysis of the pseudogenes in chromosomes 21 and 22 (англ.) // Genome Res (англ.) (рус. : journal. — 2002. — Vol. 12, no. 2. — P. 272 — 80.
- ↑ Harrison P., Gerstein M. Studying genomes through the aeons: protein families, pseudogenes and proteome evolution (англ.) // J Mol Biol (англ.) (рус. : journal. — 2002. — Vol. 318, no. 5. — P. 1155 — 74.
- ↑ Soller M. Molecular fossils in the human genome: identification and analysis of the pseudogenes in chromosomes 21 and 22 (англ.) // Cell Mol Life Sci : journal. — 2006. — Vol. 63, no. 7—9. — P. 796 — 819. (недоступная ссылка)
- ↑ Michalak P. [www.blackwell-synergy.com/doi/abs/10.1111/j.1420-9101.2006.01141.x RNA world — the dark matter of evolutionary genomics] (англ.) : journal. — 2006. — Vol. 19, no. 6. — P. 1768 — 74. Архивировано 28 января 2019 года.
- ↑ Cheng J., Kapranov P., Drenkow J., Dike S., Brubaker S et al. RNA world — the dark matter of evolutionary genomics (англ.) : journal. — 2005. — Vol. 308. — P. 1149 — 54.
- ↑ Mattick J. S. RNA regulation: a new genetics? (англ.) // Nat Rev Genet : journal. — 2004. — Vol. 5. — P. 316—323.
- ↑ Albà M. Replicative DNA polymerases (англ.) // Genome Biol (англ.) (рус. : journal. — 2001. — Vol. 2, no. 1. — P. REVIEWS3002.
- ↑ Sandman K., Pereira S., Reeve J. Diversity of prokaryotic chromosomal proteins and the origin of the nucleosome (англ.) // Cell Mol Life Sci : journal. — 1998. — Vol. 54, no. 12. — P. 1350 — 64.
- ↑ Dame R. T. The role of nucleoid-associated proteins in the organization and compaction of bacterial chromatin (англ.) // Microbiology (англ.) (рус. : journal. — Microbiology Society (англ.) (рус., 2005. — Vol. 56, no. 4. — P. 858—870. — PMID 15853876.
- ↑ Luger K., Mäder A., Richmond R., Sargent D., Richmond T. Crystal structure of the nucleosome core particle at 2.8 A resolution (англ.) // Nature : journal. — 1997. — Vol. 389, no. 6648. — P. 251 — 60.
- ↑ Jenuwein T., Allis C. Translating the histone code (англ.) // Science. — 2001. — Vol. 293, no. 5532. — P. 1074 — 80.
- ↑ Ito T. Nucleosome assembly and remodelling (неопр.) // Curr Top Microbiol Immunol. — Т. 274. — С. 1 — 22.
- ↑ Thomas J. HMG1 and 2: architectural DNA-binding proteins (англ.) // Biochem Soc Trans (англ.) (рус. : journal. — 2001. — Vol. 29, no. Pt 4. — P. 395 — 401.
- ↑ Grosschedl R., Giese K., Pagel J. HMG domain proteins: architectural elements in the assembly of nucleoprotein structures (англ.) // Trends Genet (англ.) (рус. : journal. — 1994. — Vol. 10, no. 3. — P. 94—100.
- ↑ Iftode C., Daniely Y., Borowiec J. Replication protein A (RPA): the eukaryotic SSB (англ.) // Crit Rev Biochem Mol Biol (англ.) (рус. : journal. — 1999. — Vol. 34, no. 3. — P. 141 — 80.
- ↑ Myers L., Kornberg R. Mediator of transcriptional regulation (англ.) // Annu Rev Biochem (англ.) (рус. : journal. — Vol. 69. — P. 729 — 49.
- ↑ Spiegelman B., Heinrich R. Biological control through regulated transcriptional coactivators (англ.) // Cell : journal. — Cell Press, 2004. — Vol. 119, no. 2. — P. 157—167.
- ↑ Li Z., Van Calcar S., Qu C., Cavenee W., Zhang M., Ren B. A global transcriptional regulatory role for c-Myc in Burkitt’s lymphoma cells (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2003. — Vol. 100, no. 14. — P. 8164 — 9.
- ↑ Schoeffler A., Berger J. Recent advances in understanding structure-function relationships in the type II topoisomerase mechanism (англ.) // Biochem Soc Trans (англ.) (рус. : journal. — 2005. — Vol. 33, no. Pt 6. — P. 1465 — 70.
- ↑ Tuteja N., Tuteja R. [www.blackwell-synergy.com/links/doi/10.1111/j.1432-1033.2004.04094.x Unraveling DNA helicases. Motif, structure, mechanism and function] (англ.) // Eur J Biochem (англ.) (рус. : journal. — 2004. — Vol. 271, no. 10. — P. 1849—1863. (недоступная ссылка)
- ↑ Bickle T., Krüger D. Biology of DNA restriction (англ.) // Microbiology and Molecular Biology Reviews (англ.) (рус. : journal. — American Society for Microbiology (англ.) (рус., 1993. — Vol. 57, no. 2. — P. 434 — 50.
- ↑ Joyce C., Steitz T. Polymerase structures and function: variations on a theme? (англ.) // American Society for Microbiology (англ.) (рус. : journal. — 1995. — Vol. 177, no. 22. — P. 6321 — 9.
- ↑ Hubscher U., Maga G., Spadari S. Eukaryotic DNA polymerases (англ.) // Annu Rev Biochem (англ.) (рус. : journal. — Vol. 71. — P. 133 — 63.
- ↑ Johnson A., O’Donnell M. Cellular DNA replicases: components and dynamics at the replication fork (англ.) // Annu Rev Biochem (англ.) (рус. : journal. — Vol. 74. — P. 283 — 315.
- ↑ Tarrago-Litvak L., Andréola M., Nevinsky G., Sarih-Cottin L., Litvak S. The reverse transcriptase of HIV-1: from enzymology to therapeutic intervention (англ.) // The FASEB Journal (англ.) (рус. : journal. — Federation of American Societies for Experimental Biology (англ.) (рус., 1994. — Vol. 8, no. 8. — P. 497—503.
- ↑ Martinez E. Multi-protein complexes in eukaryotic gene transcription (неопр.) // Plant Mol Biol. — 2002. — Т. 50, № 6. — С. 925 — 47.
- ↑ Cremer T., Cremer C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells (англ.) // Nat Rev Genet : journal. — 2001. — Vol. 2, no. 4. — P. 292—301.
- ↑ Pál C., Papp B., Lercher M. An integrated view of protein evolution (англ.) // Nat Rev Genet : journal. — 2006. — Vol. 7, no. 5. — P. 337 — 48.
- ↑ O’Driscoll M., Jeggo P. The role of double-strand break repair — insights from human genetics (англ.) // Nat Rev Genet : journal. — 2006. — Vol. 7, no. 1. — P. 45 — 54.
- ↑ Dickman M., Ingleston S., Sedelnikova S., Rafferty J., Lloyd R., Grasby J., Hornby D. The RuvABC resolvasome (англ.) // Eur J Biochem (англ.) (рус. : journal. — 2002. — Vol. 269, no. 22. — P. 5492 — 501.
- ↑ Joyce G. The antiquity of RNA-based evolution (англ.) // Nature. — 2002. — Vol. 418, no. 6894. — P. 214 — 21.
- ↑ Orgel L. Prebiotic chemistry and the origin of the RNA world (англ.) // Crit Rev Biochem Mol Biol (англ.) (рус. : journal. — Vol. 39, no. 2. — P. 99 — 123. Архивировано 28 июня 2007 года.
- ↑ Davenport R. Ribozymes. Making copies in the RNA world (англ.) // Science. — 2001. — Vol. 292, no. 5520. — P. 1278. — PMID 11360970.
- ↑ Szathmáry E. What is the optimum size for the genetic alphabet? (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1992. — Vol. 89, no. 7. — P. 2614 — 8. — PMID 1372984.
- ↑ Vreeland R., Rosenzweig W., Powers D. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal (англ.) // Nature : journal. — 2000. — Vol. 407, no. 6806. — P. 897 — 900.
- ↑ Hebsgaard M., Phillips M., Willerslev E. Geologically ancient DNA: fact or artefact? (англ.) // Trends Microbiol (англ.) (рус. : journal. — 2005. — Vol. 13, no. 5. — P. 212 — 20.
- ↑ Nickle D., Learn G., Rain M., Mullins J., Mittler J. Curiously modern DNA for a «250 million-year-old» bacterium (англ.) // J Mol Evol (англ.) (рус. : journal. — 2002. — Vol. 54, no. 1. — P. 134 — 7.
Литература
- Альбертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки в 3-х томах. — М.: Мир, 1994. — 1558 с. — ISBN 5-03-001986-3.
- Докинз Р. Эгоистичный ген. — М.: Мир, 1993. — 318 с. — ISBN 5-03-002531-6.
- История биологии с начала XX века до наших дней. — М.: Наука, 1975. — 660 с.
- Льюин Б. Гены. — М.: Мир, 1987. — 544 с.
- Пташне М. Переключение генов. Регуляция генной активности и фаг лямбда. — М.: Мир, 1989. — 160 с. Все форумы > Книга «переключение генов» М. Пташне.
- Уотсон Дж. Д. Двойная спираль: воспоминания об открытии структуры ДНК. — М.: Мир, 1969. — 152 с.
Ссылки
- Методы выделения и исследования ДНК.
- Веб-адреса молекулярно-биологических журналов.
- Международная база данных — последовательности ДНК из разных организмов (англ.).
- Веб-сайт Сэнгеровского Института одного из мировых лидеров в области определения последовательностей ДНК и их анализа (англ.).
- Software for forensic DNA Typing — eQMS::DNA.
| Эта статья входит в число избранных статей русскоязычного раздела Википедии. |
Некоторые внешние ссылки в этой статье ведут на сайты, занесённые в спам-лист.Эти сайты могут нарушать авторские права, быть признаны неавторитетными источниками или по другим причинам быть запрещены в Википедии. Редакторам следует заменить такие ссылки ссылками на соответствующие правилам сайты или библиографическими ссылками на печатные источники либо удалить их (возможно, вместе с подтверждаемым ими содержимым).Список проблемных доменов
|
